研究テーマ
知りたいことと見つけたこと。
概要
私たちの研究グループでは、⽣物種どうしが互いにおよぼす相互作⽤と、そうした種間関係をつうじて進化した⽣物のさまざまな性質を理解することをめざしています。得意な研究対象は、被⼦植物の花と蜜や花粉を求めて花を訪れるポリネーター(花粉媒介動物)の相互作⽤系です。
ひとくちに花とポリネーターの相互作用と言っても、そこにひそむ疑問は数かぎりなく、私たちが手がけてきたテーマも決して1つや2つではありません。そうした多岐にわたる疑問に答えるため、私たちは、野外調査を基本としつつも、ときには室内実験、あるいはコンピュータ・シミュレーションを含む数理モデルなど、その都度ふさわしいと思われるさまざまな手法を採用してきました。昔ある先輩がおっしゃった言葉を借りれば「生態学とは手法ではなく、考え方の学問だ」というのが、私たちの研究でも日々実感されることです。言いかえれば、つねに疑問やアイデアを主体に据え、手法へのこだわりを捨てることが、私たちの研究グループ(あるいは多くのマクロ生物学)の特徴と言えるかもしれません。
また生態学の研究では、しばしば専門分野を越えた広い知識や技術が必要とされます。そのため私たちは、内外のさまざまな研究者たちと共同研究を行ってきました。まだ計画段階の研究や、未発表データの論文執筆まで含めると、現在は、主に以下の方々と共同研究を進めています。
矢原徹一, James Thomson(カナダ), 川口利奈, 鈴木美季, 船本大智, 川窪伸光, 小沼明弘, Andreas Jürgens(ドイツ), Klaus Lunau(ドイツ), Mani Shresta (オーストラリア・他多数;花色の進化に関する国際研究プロジェクト)(敬称略)

主な研究テーマ
他にも、同種個体間の関係、⾷う⾷われる関係、あるいは⼈間活動と動植物の関わりなど、広く「⽣物」どうしの相互作⽤に関⼼があります。以下とは異なるテーマに取り組みたい⼈も、ぜひご相談ください。なお、最近の大橋の研究内容について知りたい方は、2021年6月に大学院の授業用にやさしく解説したセミナーの動画(前編・中編・後編;ぜんぶで42分ていど)をご覧ください。
キーワード:種間相互作用、花生態学、送受粉、採餌行動、学習、認知、適応進化、拡散共進化
①吸蜜をむずかしくする花
同じ株に長くとどまるのは、ポリネーターにとっては楽な食事法ですが、花にとっては自家受粉(=究極の近親交配)をふやす大変迷惑な行動です。吸蜜の手間がかかる複雑なかたちの花は、こうしたポリネーターの長期滞在をふせぐ機能をもつことがわかりました(Ohashi 2001, 下の解説)。
ポリネーターが訪花に費やすコストは、移動と吸蜜にかかる手間の合計ですから、吸蜜の手間がふえると、株を移動して新たな花を訪れることのコストは、今いる株で次の花を訪れるコストとほぼ等しくなります。よってポリネーターは、移動コストよりも同じ花をくり返し訪れるムダをさけるため、ひんぱんに株を移動するようになるのです。
自然界にみられる複雑なかたちの花が、自家受粉の回避という恩恵をどれだけ受けているのか、そのことが植物集団の存続にどんな影響をおよぼすのか、といった点にも興味がもたれます。
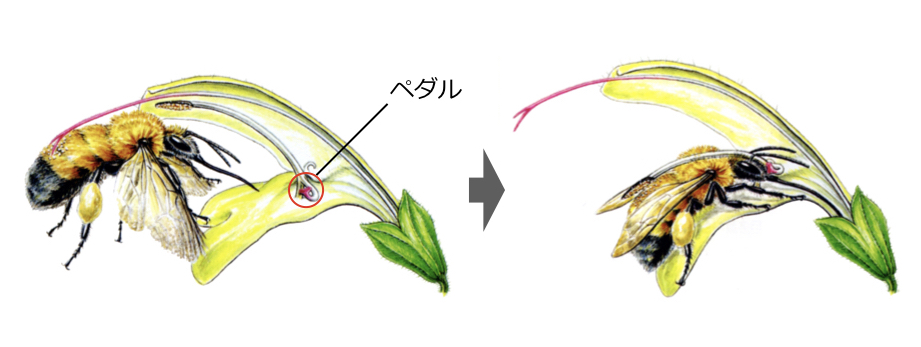
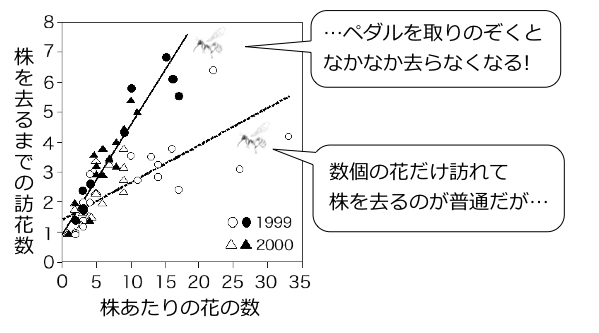
トラマルハナバチは長い舌をもち、細長いキバナアキギリの花の奥から蜜を吸うことができる。しかしキバナアキギリの花はペダル状の構造が入り口をふさいでおり、ハチが蜜を吸うには、このペダルを頭で押し上げる手間がかかる(上段 『フィールド・ウォッチング 秋の野山を歩く[北隆館]』より)。実験的にこのペダルを取りのぞいたキバナアキギリ集団をつくると、訪れたハチが株に長くとどまるようになった(下段 Ohashi 2001より改変)。
②学習するポリネーター
温帯の主要なポリネーター=マルハナバチの働きバチ(雌)は、効率良く蜜や花粉をあつめる能力に長けています。彼女らは、経験を積むほど「お気に入りの株」の位置をおぼえたり、それらの株を「お気に入りのルート」に沿って何度も訪れたりするようになります(Makino, Ohashi & Sakai 2007, 下の解説)。
経験を積んだトラップライナーは、何度も戻ってくるだけでなく、すぐに同じ株へ戻ることもないため、多くの花粉を遠くに届けてくれるありがたい存在です(Ohashi & Thomson 2009)。しかしその反面、1つ1つの株をおぼえているため、たとえば花がこっそり蜜を出すのを止めたりすると、すぐに見やぶって戻ってこなくなる、というやっかいな面もあります。
いわば、花にとってマルハナバチは、「ありがたいけれど注文の多い固定客」なのです。花は、どんな手を使って、彼女らをうまくポリネーターとして利用しているのでしょうか?
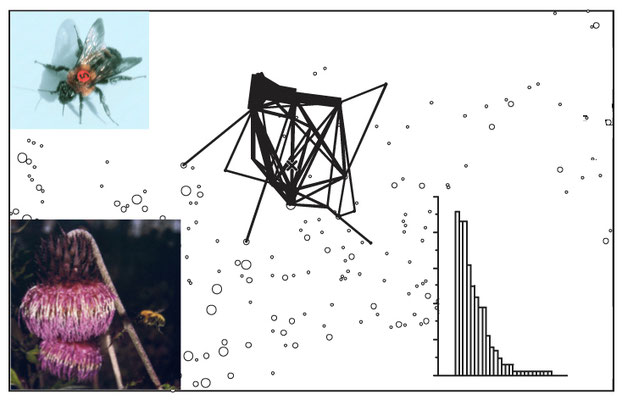
フジアザミの自生集団を訪れるマルハナバチに背番号をつけ、株から株への移動を追跡した。すると、個体はしばしばきまった株を、一定のルートに沿って巡回することがわかった(Makino, Ohashi & Sakai 2007)。また、同じルートをたどる傾向が強い個体ほど、花数が多い株をルートに組みこんでいた。これは、マルハナバチが餌あつめの経験を積むほど質の良い株をおぼえ、自分だけのお気に入りのルート=トラップラインを学習するからだ(Ohashi et al. 2007)。
トラップライナーは、隣りあう株を何度も行ったり来たりするといったムダな動きをしない。コンピュータ・シミュレーションで検討したところ、トラップライナーのこうした動きは、より多くの花粉を遠くまで届けるという、植物にとってプラスの効果をもたらすことがわかった(Ohashi & Thomson 2009)。
③看板を塗りかえる花
自然界には、ポリネーターを呼ぶ大切な看板であるはずの花の色を、咲き終わるまえにがらりと変えてしまう花色変化植物が存在します(Ohashi et al. 2015, 下の解説)。色を変えるのは、蜜も花粉も出さなくなった古い花です。
花色変化は、古い花を維持して株を目立たせ、見かけの派手さにだまされやすいポリネーターをおびきよせつつ、色を変えて報酬の有無を知らせ、学習経験を積んだマルハナバチのような「ありがたいけれど注文の多い固定客」(②参照)も確保する、よくばりな戦略と考えられます。
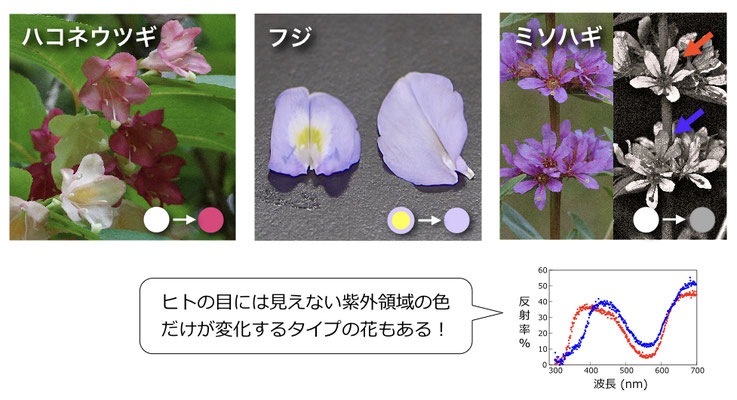
花が咲いてからしぼむまでにどれくらい色が変化するのか、219種の波長スペクトルを調べると、そのうち約2割が顕著な花色変化を示し、さらにその1/4は、ヒトには見えない紫外領域の色だけが大きく変化することがわかった (Ohashi et al. 2015)。色変化は花びら全体で起こる種もあれば、一部の箇所だけで起こる種もある。その理由はまだよくわかっていない。
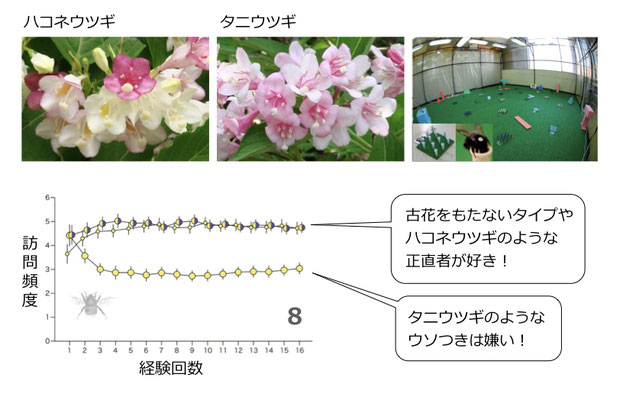
花色変化するハコネウツギ(左)と、色を変えずに古い花を維持するタニウツギ(中央)を植物園内でくらべたところ、前者の方がハナバチに好まれていた(Suzuki & Ohashi 2014)。室内実験(右)でも、ハコネウツギのようなタイプは経験量によらず常にマルハナバチに好まれる反面、タニウツギのようなタイプは、経験を積んだマルハナバチには嫌われてしまうことが示された(下 Makino & Ohashi 2017より改変)。
④きまった時刻に開く花
何日間も咲きつづける花にとって、最初につぼみを開く時刻はさほど重要な意味をもたないように思えます。ところが実際は、ほぼきまった時刻に開く花が多くみられます。しかもその時刻は植物種ごとに、朝、昼、夕、夜とさまざまです。これにはどんな意味があるのでしょう?

夕咲きの花、ツリガネニンジンの受粉は夜行性のガによっておこなわれます。翌日の昼にはハナバチやハナアブも訪れますが、受粉の効率は劣るようです(Funamoto & Ohashi 2017, 下の解説)。よって夕方に花を開けば、受粉の効率が劣るハナバチやハナアブの訪花の順番を後にできるため、花粉や蜜、柱頭のムダをさけ、昼と夜のポリネーターを最も効果的に活用できると考えられます。
他の花の開花時刻にも同様の説明があてはまるのか、活動時間が異なるポリネーターの習性の差は花粉の受け渡しの効率にどんな影響をおよぼすのか、といった点にも興味がもたれます。

ツリガネニンジンの花は、日没の直後に開花する。また、花は数日間にわたり咲きつづけるものの、蜜を分泌するのは夜間のみである。袋がけ実験の結果、受粉はもっぱら夜に訪れるガのなかま(1, 2)によっておこなわれ、翌日の昼に訪れるハナバチ(3)やハナアブ(4)は、花粉や蜜をムダに消費するばかりで受粉にあまり役立たないことがわかった(Funamoto & Ohashi 2017)。ツリガネニンジンの夕咲きという性質は、効率の高いポリネーターであるガを優先的に利用する一方で、夜の間にガが来なかった場合の「保険」として、昼のポリネーターによる訪花の可能性も残しておくための戦略と考えられる。
予想してその結果が近かったら、その考えが正しかったかもしれないからうれしい。
しかし結果が違っていたら、それまでに考えもしなかった
ほかの考えに近いからうれしいのだ。
そのほうがずっといい。
— B. ハインリッチ